



一、Increase the sensitivity of the reaction system:
1. Isolate high quality RNA:
Successful cDNA synthesis comes from high-quality RNA. High-quality RNA should at least be full-length and free of reverse transcriptase inhibitors such as EDTA or SDS. The quality of RNA determines the maximum amount of sequence information you can transcribe into cDNA. A common RNA purification method is a one-step method using guanidine isothiocyanate/acid phenol. To prevent contamination by trace amounts of RNase, RNA isolated from RNase-rich samples (such as pancreas) needs to be stored in formaldehyde to preserve high-quality RNA, especially for long-term storage. The RNA extracted from rat liver was basically degraded after being stored in water for one week, while the RNA extracted from rat spleen remained stable after being stored in water for 3 years. In addition, transcripts longer than 4 kb are more sensitive to degradation by trace RNases than small transcripts. To increase the stability of stored RNA samples, RNA can be dissolved in deionized formamide and stored at -70°C. Formamide used to preserve RNA must be free of RNA-degrading debris. RNA from pancreas can be preserved in formamide for at least one year. When preparing to use RNA, you can use the following method to precipitate RNA: add NaCl to 0.2M and 4 times the volume of ethanol, place at room temperature for 3-5 minutes, and centrifuge at 10,000×g for 5 minutes.
2. Use the RNaseH-inactive (RNaseH-) reverse transcriptase:
RNase inhibitors are often added to reverse transcription reactions to increase the length and yield of cDNA synthesis. RNase inhibitors should be added during the first-strand synthesis reaction in the presence of a buffer and a reducing agent (such as DTT), because the process prior to cDNA synthesis denatures the inhibitor, thereby releasing bound RNase that can degrade RNA. Protein RNase inhibitors only prevent the degradation of RNA by RNase A, B, C, and do not prevent RNase on the skin, so be careful not to introduce RNase from your fingers despite the use of these inhibitors.
Reverse transcriptase catalyzes the conversion of RNA to cDNA. Both M-MLV and AMV have endogenous RNaseH activity in addition to their own polymerase activity. RNaseH activity and polymerase activity compete with each other for the hybrid strand formed between the RNA template and the DNA primer or cDNA extension strand, and degrade the RNA strand in the RNA:DNA complex. The RNA template degraded by RNaseH activity can no longer serve as an effective substrate for cDNA synthesis, which reduces the yield and length of cDNA synthesis. Therefore, it would be beneficial to eliminate or greatly reduce the RNaseH activity of reverse transcriptase.。
SuperScript Ⅱ reverse transcriptase, RNaseH- MMLV reverse transcriptase and thermoScript reverse transcriptase, RNaseH- AMV, can obtain more amount and more full-length cDNA than MMLV and AMV. RT-PCR sensitivity will be affected by the amount of cDNA synthesis. ThermoScript is much more sensitive than AMV. The size of RT-PCR products is limited by the ability of reverse transcriptase to synthesize cDNA, especially when cloning larger cDNAs. Compared with MMLV, SuperScripⅡ significantly increased the yield of long RT-PCR products. The RNaseH-reverse transcriptase also has increased thermostability, so the reaction can be performed at temperatures higher than the normal 37-42°C. Under the suggested synthesis conditions, use oligo(dT) primer and 10 μCi of [α-P]dCTP. The total yield of first strand was calculated using the TCA precipitation method. Full-length cDNA was analyzed using size-sorted bands excised and counted on an alkaline agarose gel.
3. Raise the incubation temperature for reverse transcription:
A higher incubation temperature helps to open the RNA secondary structure, increasing the yield of the reaction. For most RNA templates, incubating the RNA and primers at 65°C without buffer or salt, followed by rapid cooling on ice will eliminate most secondary structures and allow primers to bind. However, some templates still have secondary structures, even after heat denaturation. Amplification of these difficult templates can be performed using ThermoScript Reverse Transcriptase and placing the reverse transcription reaction at a higher temperature to improve amplification. Higher incubation temperatures can also increase specificity, especially when gene-specific primers (GSP) are used for cDNA synthesis (see Chapter 3). If using GSP, ensure that the Tm of the primers is the same as the expected incubation temperature. Do not use oligo(dT) and random primers above 60°C. Random primers require incubation at 25°C for 10 minutes before increasing to 60°C. In addition to using a higher reverse transcription temperature, specificity can also be improved by directly transferring the RNA/primer mix from the 65°C denaturation temperature to the reverse transcription incubation temperature and adding a pre-warmed 2× reaction mixture (cDNA hot-start synthesis) . This approach helps prevent the intermolecular base pairing that occurs at lower temperatures. The multiple temperature switching required for RT-PCR can be simplified by using a thermal cycler.
Tth thermostable polymerase acts as a DNA polymerase in the presence of Mg2+ and as an RNA polymerase in the presence of Mn2+. It can be kept warm at a maximum temperature of 65°C. However, the presence of Mn2+ during PCR reduces fidelity, which makes Tth polymerase less suitable for high-precision amplification, such as cloning of cDNA. In addition, Tth has low reverse transcription efficiency, which reduces sensitivity, and, since reverse transcription and PCR can be performed with a single enzyme, control reactions without reverse transcription cannot be used to compare cDNA amplification products with contaminating genomic DNA. The amplification products were separated.
4. Additives that promote reverse transcription:
Additives including glycerol and DMSO are added to the first-strand synthesis reaction, which can reduce the stability of the nucleic acid double-strand and untie the secondary structure of RNA. Up to 20% glycerol or 10% DMSO can be added without affecting SuperScript II or MMLV activity. AMV can also tolerate up to 20% glycerol without loss of activity. In order to maximize the sensitivity of RT-PCR in the SuperScriptⅡ reverse transcription reaction, 10% glycerol can be added and incubated at 45°C. If 1/10 of the reverse transcription reaction product is added to PCR, then the concentration of glycerol in the amplification reaction is 0.4%, which is not enough to inhibit PCR.
5. RNaseH treat:
Treatment of cDNA synthesis reactions with RNaseH prior to PCR can increase sensitivity. For some templates, it is thought that RNA in the cDNA synthesis reaction prevents the binding of amplification products, in which case RNaseH treatment can increase sensitivity. Generally, RNaseH treatment is necessary when amplifying longer full-length cDNA target templates, such as low-copy tuberous scherosis II. For this difficult template, RNaseH treatment enhanced the signal produced by SuperScript II or AMV-synthesized cDNA. For most RT-PCR reactions, RNaseH treatment is optional, because the PCR denaturation step at 95°C generally hydrolyzes the RNA in the RNA:DNA complex.
6. Improvement of Small RNA Detection Method:
RT-PCR is especially challenging when only small amounts of RNA are available. Glycogen added as a carrier during RNA isolation helps to increase the yield of small samples. RNase-free glycogen can be added at the same time as adding Trizol. Glycogen is water soluble and can be kept in the aqueous phase with RNA to aid subsequent precipitation. For samples less than 50 mg of tissue or 106 cultured cells, the recommended concentration of RNase-free glycogen is 250 μg/ml.
Adding acetylated BSA to the reverse transcription reaction using SuperScript II can increase the sensitivity, and for small amounts of RNA, reducing the amount of SuperScript II and adding 40 units of RNaseOut nuclease inhibitor can increase the level of detection. If glycogen is used in the RNA isolation process, it is still recommended to add BSA or RNase inhibitor when using SuperScript II for reverse transcription reaction.
二、Increase RT-PCR specificity
1. CND Asynthesis:
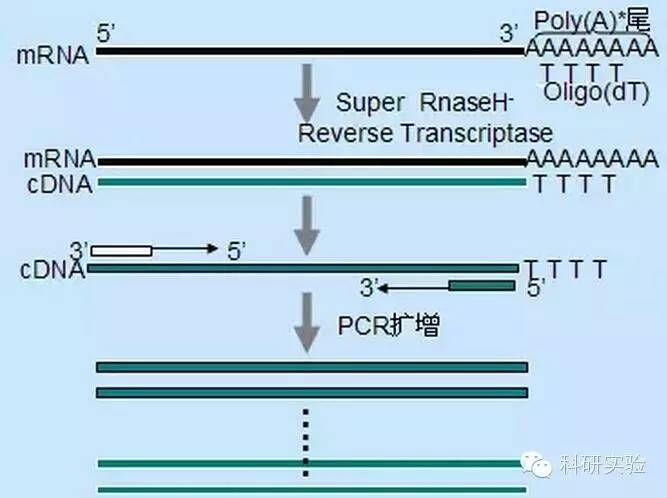
First-strand cDNA synthesis can be initiated using three different methods, the relative specificity of which affects the amount and type of cDNA synthesized.
The random primer method was the least specific of the three methods. Primers anneal at multiple sites throughout the transcript, generating short, partial-length cDNAs. This method is frequently used to obtain 5′ end sequences and to obtain cDNA from RNA templates with regions of secondary structure or with termination sites that cannot be replicated by reverse transcriptase. To obtain the longest cDNA, the ratio of primers to RNA in each RNA sample needs to be determined empirically. The starting concentration of random primers ranged from 50 to 250 ng per 20 μl reaction. Since cDNA synthesized from total RNA using random primers is primarily ribosomal RNA, poly(A)+RNA is generally chosen as the template.
Oligo(dT) primers are more specific than random primers. It hybridizes to the poly(A) tail found at the 3′ end of most eukaryotic mRNAs. Because poly(A)+ RNA is approximately 1% to 2% of total RNA, the amount and complexity of cDNA is much less than with random primers. Because of its high specificity, oligo(dT) generally does not require optimization of the ratio of RNA to primers and poly(A)+ selection. It is recommended to use 0.5μg oligo(dT) per 20μl reaction system. oligo(dT)12-18 is suitable for most RT-PCR. The ThermoScript RT-PCR System offers oligo(dT)20 because of its better thermal stability for higher incubation temperatures.
Gene specific primers (GSP) are the most specific primers for the reverse transcription step. GSP is an antisense oligonucleotide that can specifically hybridize to the RNA target sequence, unlike random primers or oligo(dT), which anneal to all RNAs. The same rules used to design PCR primers apply to the design of GSP in reverse transcription reactions. The GSP can be the same sequence as the amplification primer that anneals to the 3′-most end of the mRNA, or the GSP can be designed to anneal downstream of the reverse amplification primer. For some amplified subjects, more than one antisense primer needs to be designed for successful RT-PCR because the secondary structure of the target RNA may prevent primer binding. It is recommended to use 1 pmol antisense GSP in a 20 μl first-strand synthesis reaction.
2. Raise the incubation temperature for reverse transcription:
In order to take full advantage of the full advantage of GSP specificity, a reverse transcriptase with higher thermostability should be used. Thermostable reverse transcriptases can be incubated at higher temperatures to increase reaction stringency. For example, if a GSP anneals at 55°C, the specificity of the GSP will not be fully utilized if AMV or M-MLV is used for reverse transcription at a low stringency of 37°C. However, SuperScript II and ThermoScript can be reacted at 50°C or higher, which will eliminate non-specific products generated at lower temperatures. For maximum specificity, the RNA/primer mix can be transferred directly from the 65°C denaturation temperature to the reverse transcription incubation temperature and added to a pre-warmed 2× reaction mix (cDNA synthesis hot start). This helps prevent intermolecular base pairing at low temperatures. The multiple temperature transitions required for RT-PCR can be simplified by using a thermal cycler.
3. Reduces genomic DNA contamination:
A potential difficulty encountered with RT-PCR is the contamination of genomic DNA in the RNA. Using a good RNA isolation method, such as Trizol Reagent, will reduce the amount of genomic DNA contaminating the RNA preparation. To avoid products derived from genomic DNA, RNA can be treated with amplification-grade DNase I to remove contaminating DNA prior to reverse transcription. The DNase I digestion was terminated by incubating the samples in 2.0 mM EDTA for 10 minutes at 65°C. EDTA can chelate magnesium ions, preventing magnesium ion-dependent RNA hydrolysis at high temperatures.
In order to separate amplified cDNA from contaminating genomic DNA amplification products, primers can be designed that each anneal to separate exons. PCR products derived from cDNA will be shorter than those derived from contaminated genomic DNA. In addition, a control experiment without reverse transcription was performed on each RNA template to determine whether a given fragment was derived from genomic DNA or cDNA. The PCR product obtained without reverse transcription is derived from the genome.
Post time: May-16-2023



